O desenvolvimento embrionário O zigoto é portador do material genético fornecido pelo espermatozóide e pelo óvulo. Um vez formado o zigoto irá se dividir muitas vezes por mitose até originar um novo indivíduo. Assim, todas as células que formam o corpo de um indivíduo possuem o mesmo patrimônio genético que existia no zigoto. Apesar disso, ao longo do desenvolvimento embrionário as células passam por um processo de diferenciação celular em que alguns genes são “ativados” e outros são “desativados”, sendo que somente os “ativados” coordenam as funções das células. Surgem dessa maneira tipos celulares com formatos e funções distintos, que se organizam em tecidos. Conjuntos de tecidos reunidos formam os órgãos. Os grupos de órgãos formam os sistemas que, por sua vez, formam o organismo. Células – tecidos – órgãos – sistemas – organismos A ciência que estuda esse processo de desenvolvimento do indivíduo a partir do zigoto é a Embriologia.  Fases do desenvolvimento embrionário Os animais apresentam grande diversidade de desenvolvimento embrionário, mas, de modo geral, em praticamente todos ocorrem três fases consecutivas: segmentação, gastrulação e organogênese. Na segmentação, mesmo com o aumento do número de células, praticamente não há aumento do volume total do embrião, pois as divisões celulares são muito rápidas e as células não têm tempo para crescer. Na fase seguinte, que é a gastrulação, o aumento do número de células é acompanhada do aumento do volume total. Inicia-se nessa fase a diferenciação celular, ocorrendo a formação dos folhetos germinativos ou folhetos embrionários, que darão origem aos tecidos do indivíduo. No estágio seguinte, que é a organogênese, ocorre a diferenciação dos órgãos. Vamos analisar cada uma dessas fases para os animais em geral e depois comentar o desenvolvimento embrionário humano. |
Segmentação
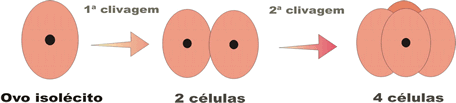
As divisões que ocorrem durante a segmentação denominam-se clivagens, e as células que se formam são chamadas blastômeros.
No Reino Animal, a diferença na quantidade e na distribuição do vitelo no ovo determina diferenças na segmentação, menor a velocidade de divisão. Em função disso, podemos considerar dois tipos básicos de segmentação:
- holoblástica ou total que ocorre no zigoto todo;
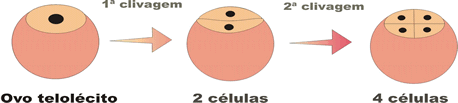
- meroblástica ou parcial, que ocorre só em parte do ovo.
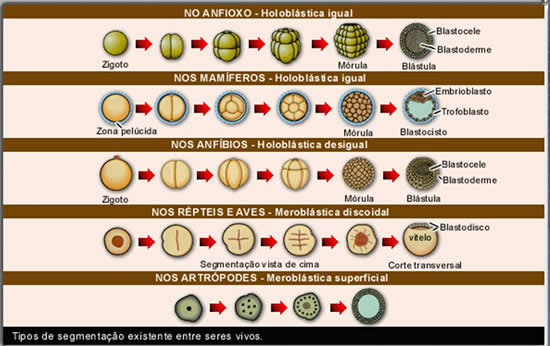
Segmentação holoblástica
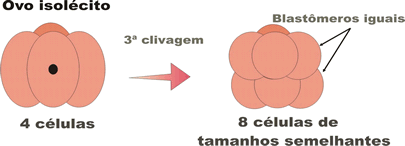
A segmentação holoblástica ocorre nos alécitos, nos isolécitos (ou oligolécitos) e nos heterolécitos, e pode ser subdividida em três tipos, com base no tamanho das células que se formam a partir da terceira clivagem (quando muda o plano de divisão celular):
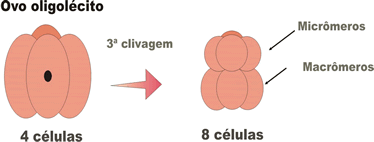
- holoblástica igual, na qual se formam, com a terceira clivagem, oito blastômeros iguais; ocorre nos ovos alécitos e em alguns oligolécitos;
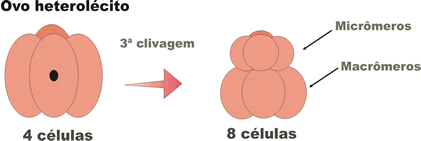
- holoblástica desigual, na qual se formam, com a terceira clivagem, blastômeros de tamanhos diferentes (quatro menores: micrômeros; e quatro maiores: macrômeros); Ocorre em todos os ovos heterolécitos e em alguns oligolécitos;
- holoblásticas subigual, um tipo de segmentação desigual em que os blastômeros não diferem muito entre si quanto ao tamanho, ocorre em alguns ovos isolécitos.
Segmentação meroblástica
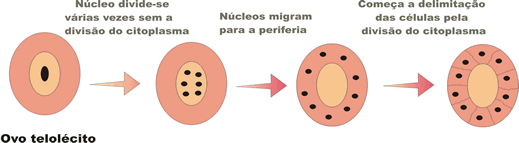
Devido à diferença na distribuição do vitelo, existem dois tipos básicos de segmentação meroblástica: a discoidal e a superficial.
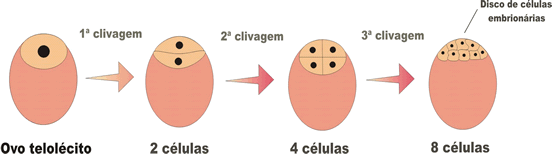
Na segmentação meroblástica discoidal, as divisões ocorrem apenas na região da cicatrícula (região da célula sem vitelo), formando-se um disco de células sobre a massa do vitelo. Esse tipo de segmentação ocorre nos ovos telolécitos.
A segmentação meroblástica superficial ocorre nos ovos centrolécitos. As células embrionárias ficam dispostas na superfície do ovo.
Fases da segmentação
Embora existam diferentes tipos de segmentação, eles normalmente se realizam segundo duas fases:
- mórula, em que se forma um maciço celular com poucas células;
- blástula, em que é aumentado o número de células e se forma uma cavidade interna cheia de líquido.

A cavidade central que se observa na blástula recebe o nome de blastocele (cele = cavidade) e é cheia de líquido sintetizado pelas células que formam os seus limites.
Nos ovos isolécitos e nos heterolécitos a blastocele é bem desenvolvida.
Na blástula originada da segmentação de ovos telolécitos, não se observa a verdadeira blastocele (cele = cavidade) e é cheia de líquido sintetizado pelas células que formam os seus limites.
Nos ovos isolécitos e nos heterolécitos a blastocele é bem desenvolvida.
Na blástula originada da segmentação de ovos telolécitos, não se observa a verdadeira blastocele, pois a cavidade formada não é inteiramente delimitada pelos blastômeros. Essa cavidade é delimitada em parte pelos blastômeros e em parte pelo vitelo. Nesse caso, a cavidade formada recebe o nome de cavidade subgerminal, que também é preenchida por líquido sintetizado pelas células. A blástula que se forma a partir da segmentação dos ovos telolécitos recebe o nome de discoblástula.
Gastrulação
Para falarmos da gastrulação, vamos tomar como exemplo o que ocorre em animais cordados, representados pelo anfioxo e pelas rãs.
Os cordados são animais que possuem notocorda, um bastonete flexível que fica no dorso do embrião. A notocorda persiste no adulto de alguns animais cordados, como é o caso do anfioxo. Nos animais vertebrados, excluindo alguns peixes, a notocorda regride totalmente ou quase totalmente e a coluna vertebral se desenvolve a partir da mesoderma.
O anfioxo é um animal de cerca de 6 cm de comprimento que vive enterrado na areia em águas rasas do ambiente marinho, deixando para fora apenas a região anterior do corpo. Esses animais têm sexos separados e a fecundação é externa. O ovo do anfioxo é oligolécito e a sua segmentação é total subigual. A gastrulação ocorre por um processo denominado invaginação dos blastômeros para o interior da blastocele, como se um dedo empurrasse a parede de uma bexiga. A blastocele se reduz e chega a desaparecer. No ponto de invaginação surge um orifício denominado blastóporo; a cavidade interna que se forma é o intestino primitivo ou arquêntero. |  |
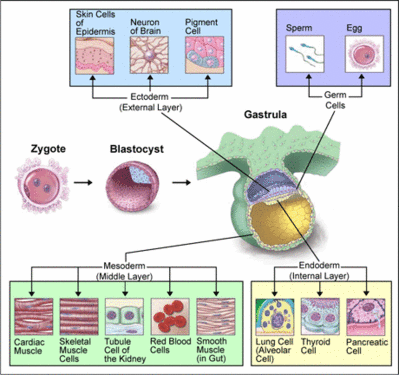
Na gastrulação, diferenciam-se os folhetos germinativos ou embrionários, que darão origem a todos os tecidos e órgãos. Esses folhetos são: ectoderma (o mais externo), mesoderma (o intermediário) e endoderma (o mais interno).
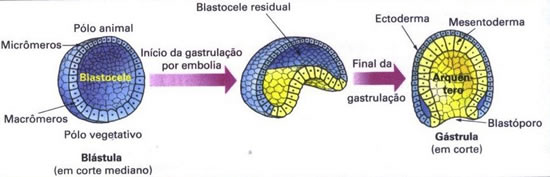
Os animais que possuem três folhetos germinativos são chamados triblásticos ou triploblásticos, como é o caso dos cordados. Existem entretanto, animais que possuem apenas dois folhetos germinativos: o ectoderma e o endoderma. Esses animais são chamados diblásticos ou diploblásticos, como e o caso dos cnidários.
O esquema acima descreve de forma simplificada a gastrulação em anfioxo. Neste caso, a camada interna que reveste diretamente o arquêntero é chamada mesentoderma e dará origem, logo a seguir ao mesoderma e ao endoderma. (Há quem considere o mesentoderma como endoderma e o mesoderma formado a partir do endoderma.)
Nas rãs a fecundação é externa, os óvulos são heterolécitos e a segmentação é total desigual. Os óvulos possuem um envoltório gelatinoso que desseca em contato com o ar. Assim, todo o desenvolvimento embrionário ocorre na água. Forma-se uma larva aquática, o girino que sofre metamorfose, originando o adulto. Fala-se, nesses casos, em desenvolvimento indireto, pois há uma fase larval. Quando a fase larval não está presente, fala-se em desenvolvimento direto. |  |
A gastrulação das rãs ocorre por invaginação e também por epibolia. Por invaginação forma-se uma fenda: o blastóporo. Por epibolia os micrômeros passam a se dividir rapidamente e acabam por recobrir os macrômeros. Os micrômeros insinuam-se primeiramente pelo lábio ventral. O blastóporo adquire o aspecto de um círculo. Os micrômeros insinuam-se para dentro da blastocele, delimitando o arquêntero. Ocorre também a diferenciação dos três folhetos germinativos: o ectoderma, o mesoderma e o endoderma.
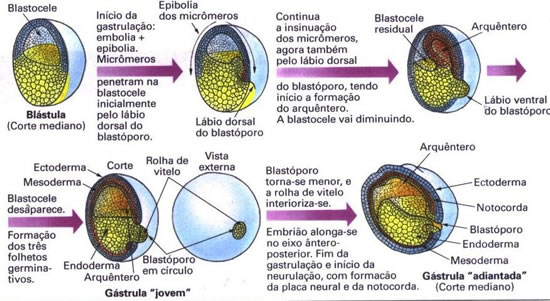
- formação dos folhetos embrionários ou germinativos, que darão origem a todos os tecidos e órgãos;
- formação do arquêntero ou intestino primitivo;
- formação do blastóporo, orifício de comunicação do arquêntero com o exterior.
Protostomados e deuterostomados
O blastóporo pode dar origem à boca ou ao ânus. Quando dá origem apenas à boca ou tanto à boca quanto ao ânus, os animais são chamados de protostômios (proto = primeiro). É o caso dos vermes, dos moluscos e dos artrópodes.
Quando o blastóporo dá origem ao ânus os animais são chamados de deuterostômios (deutero = posterior). É o caso dos equinodermos e dos cordados.

Organogênese em anfioxo
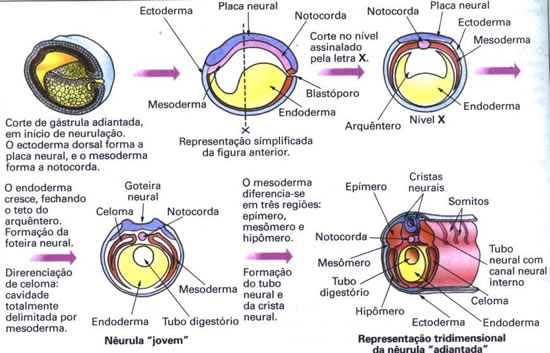
A terceira fase do desenvolvimento embrionário é a organogênese, que se caracteriza pela diferenciação de órgãos a partir dos folhetos embrionários formados na gastrulação. O esquema seguinte representa a fase inicial da organogênese: a neurulação. Após a neurulação, os folhetos embrionários, continuam a se diferenciar, originando os tecidos especializados do adulto.
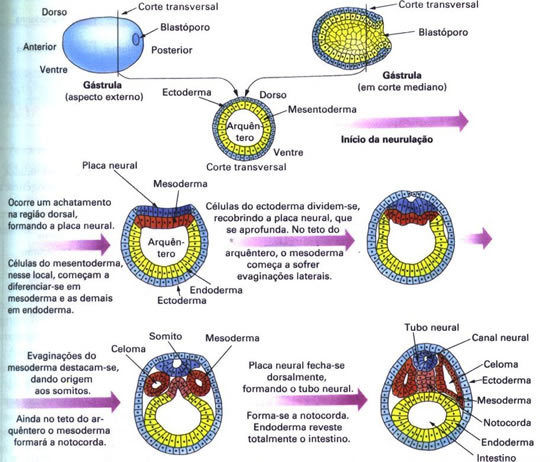
Do ectoderma diferencia-se o tubo neural, que apresenta no seu interior o canal neural. O endoderma dá origem ao tubo digestório. O mesoderma dá origem aos somitos e à notocorda. Os somitos são blocos celulares dispostos lateralmente no dorso do embrião, e a notocorda é uma estrutura maciça localizada logo abaixo do tubo neural.
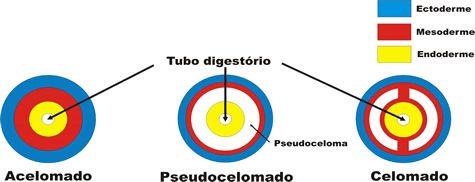
O mesoderma delimita cavidades denominadas celomas.
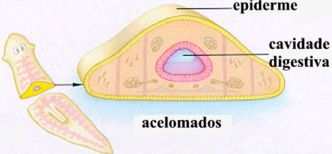
Animais celomados, acelomados e pseudocelomados
Os animais que apresentam celoma são chamados celomados. Todos os cordados são celomados , assim como os moluscos (lesmas, ostras), os anelídeos (minhocas) e os equinodermos (estrelas-do-mar, ouriços-do-mar).
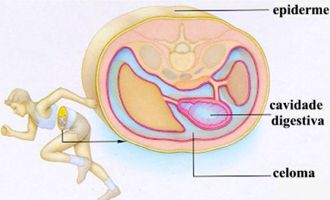
Há animais triblásticos em que a mesoderma delimita uma parte da cavidade, sendo a outra parte delimitada pela endoderma. Esses animais são chamados de pseudocelomados, pois o celoma só é verdadeiro quando é completamente revestido pelo mesoderma. É o caso dos nematódeos, cujo representante mais conhecido é a lombriga (Ascaris lumbricóides), um parasita do intestino humano.
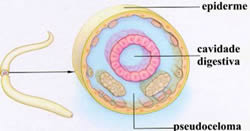
Em alguns animais a única cavidade que se forma no embrião é o arquêntero, pelo que são designados acelomados.
O esquema a seguir mostra cortes transversais esquemáticos em organismos acelomados, pseudocelomados e celomados:
Organogênese em rã
A organogênese em rã será estudada como exemplo da organogênese geral dos vertebrados. O esquema a seguir explica de forma simplificada como ocorre a fase inicial da organogênese nesses animais: a neurulação.
Alguns dos destinos finais dos folhetos embrionários nos vertebrados em geral são:
Ectoderma | Mesoderme | Endoderme |
|
|
|
Células embrionárias especiais típicas e exclusivas dos vertebrados formam a crista neural. Essas células diferenciam-se juntamente com a formação do tubo neural a partir do ectoderma do embrião. Elas ficam dispostas ao lado do tubo neural, mas posteriores à região do encéfalo.
As células da crista neural têm a capacidade de migrar pelo corpo, dando origem a diversos tipos celulares, como neurônios sensoriais do sistema nervoso periférico, células da medula da adrenal, derme da pele da cabeça e células pigmentares da pele de todo o corpo.
Considera-se hoje que o passo definitivo na origem dos vertebrados tenha sido a evolução das células da crista neural. Nenhum outro animal as possui.
Anexos embrionários: Adaptação ao Meio Terrestre
Anexos embrionários são estruturas que derivam dos folhetos germinativos do embrião mas que não fazem parte do corpo desse embrião.
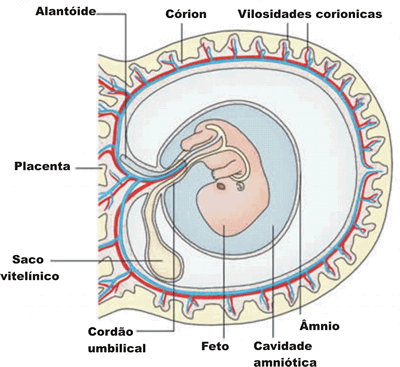
Os anexos embrionários são: vesícula vitelina (saco vitelínico), âmnio (ou bolsa amniótica), cório e alantóide.
Vesícula vitelina
Durante a evolução do grupo dos animais, os primeiros vertebrados que surgiram foram os peixes, grupo que possui como único anexo embrionário a vesícula vitelina.
Diferenciando-se a mesoderme e o tubo neural, parte dos folhetos germinativos desenvolvem-se formando uma membrana que envolve toda a gema, constituindo (membrana + gema) o saco vitelínico um anexo embrionário, que permanece ligado ao intestino do embrião. À medida que este se desenvolve, há o consumo do vitelo e, consequentemente, o saco vitelínico vai se reduzindo até desaparecer. É bem desenvolvida não somente em peixes, mas também em répteis e aves. Os mamíferos possuem vesícula vitelina reduzida, pois nesses animais como regra geral, os ovos são pobres em vitelo. A vesícula vitelina não tem, portanto, significado no processo de nutrição da maioria dos mamíferos.
Nos anfíbios, embora os ovos sejam ricos em vitelo, falta a vesícula vitelina típica. Nesses animais o vitelo encontra-se dentro de células grandes (macrômeros) não envoltas por membrana vitelina própria.
Âmnio e cório
O âmnio é uma membrana que envolve completamente o embrião, delimitando uma cavidade denominada cavidade amniótica. Essa cavidade contém o líquido amniótico, cujas funções são proteger o embrião contra choques mecânicos e dessecação. Ao final do desenvolvimento de répteis e aves, todo o líquido da cavidade amniótica foi absorvido pelo animal.
O cório ou serosa é uma membrana que envolve o embrião e todos os demais anexos embrionários. É o anexo embrionário mais externo ao corpo do embrião. Nos ovos de répteis e nos de aves, por exemplo, essa membrana fica sob a casca. Nesses animais, o cório, juntamente com o alantóide, participa dos processos de trocas gasosas entre o embrião e o meio externo.
Alantóide
A alantóide é um anexo que deriva da porção posterior do intestino do embrião. A função da alantóide nos répteis e nas aves é: transferir para o embrião as proteínas presentes na clara, transferir parte dos sais de cálcio, presentes na casca, para o embrião, que utilizará esses sais na formação de seu esqueleto, participar das trocas gasosas, o O2 passa da câmera de ar para o alantóide e deste para o embrião, enquanto o CO2 produzido percorre o caminho inverso, e armazenar excreta nitrogenada. A excreta nitrogenada eliminada por embriões desses animais é o ácido úrico, insolúvel em água e atóxico, podendo ser armazenado no interior do ovo sem contaminar o embrião.
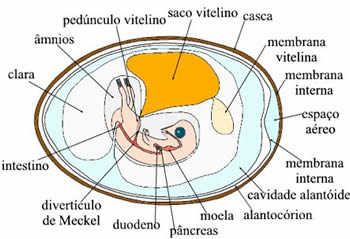
O ovo e a gema Em aves, o termo ovo pode ser empregado para todo o conjunto formado pela casca, clara e gema; ou, então, ser usado no sentido estritamente embriológico: óvulo fecundado ou zigoto. Nas galinhas, o ovócito II é liberado dos ovários e penetra no oviduto, onde poderá ser fecundado pelo espermatozóide. O desenvolvimento embrionário inicia-se no oviduto e é concluído fora do corpo da ave. O ovo em desenvolvimento, quando ainda está na porção anterior do oviduto, é envolto por um albume denso, secretado por células glandulares da parede desse órgão que serve de alimento para o embrião. Nessa região, além de glândulas, há pregas espiraladas que determinam a rotação do ovo quando ele passa pelo oviduto, Isso faz com que o albume envolva intimamente a gema, desse modo formando, de cada lado, uma corda enrolada mais opaca, denominada calaza. Posteriormente, no oviduto, é novamente adicionado albume só que menos consistente. Uma película elástica é acrescentada ao redor de toda a clara. Na porção terminal, é secretada a casca calcária porosa, formada por carbonatos e fosfatos de cálcio e magnésio. O tempo de permanência do embrião em desenvolvimento dentro do oviduto é de cerca de 24 horas, no caso das galinhas. Quando o ovo é posto, o embrião já está no final da segmentação. Após um período de incubação de 21 dias a 37,5ºC ocorre a eclosão. A gema do ovo pode corresponder ao ovócito II quando não há fecundação, ou ao embrião quando ocorre a fecundação. | 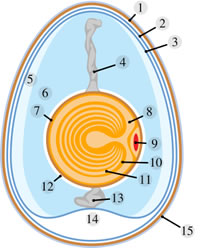 | |
| 1 - Membrana da Conquilha 4 - Calaza 6 - Clara 7- Membrana vitelínica 9 - Germen 13 - Calaza 14 - Câmara de Ar 15 - Conquilha ou casca |
Mamíferos: Surge a placenta
Na maioria dos mamíferos, o desenvolvimento embrionário ocorre no interior do corpo materno, dentro de um órgão musculoso, o útero. Excetuando os mamíferos que botam ovos (ornitorrinco e equidna), todos os demais formam a placenta, órgão constituído pela parede interna vascularizada do útero (endométrio) e por estruturas derivadas do trofoblasto ou trofoderme embrionário (nos mamíferos, nome dado à câmara mais externa de revestimento do embrião). Alimentos, oxigênio, anticorpos e hormônios passam do sangue materno para o embrionário, pela placenta, que, em troca, transfere para a mãe as excretas e o gás carbônico.
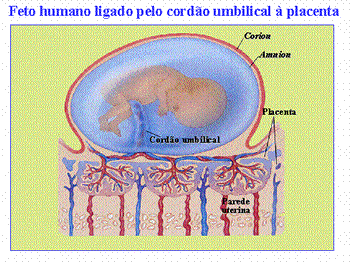
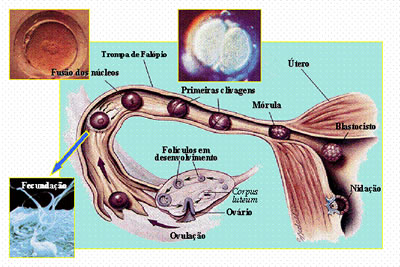
No homem, o ovo é do tipo oligolécito e a segmentação (clivagem) é total e igual, logo se formando a fase de mórula. Atingida essa fase, o embrião ingressa na cavidade uterina. No interior dessa cavidade, surge a fase correspondente a blástula, que, nos mamíferos, é denominada blastocisto. Nesse estágio, o embrião é dotado de uma camada externa de células, o trofoblasto, que envolve um aglomerado interno de células, a massa celular interna. Cabe a essa massa celular a formação do corpo do embrião, enquanto o trofoblasto será o responsável pela penetração do embrião no interior do endométrio (a camada interna da parede uterina), e pela organização da parte embrionária da placenta.
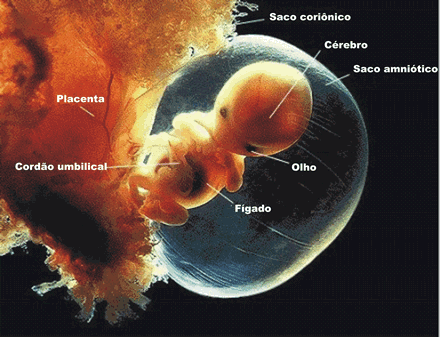
No embrião humano, o trofoblasto e a mesoderme extra-embrionária formam o cório. Esse duplo revestimento é responsável pela organização das vilosidades coriônicas, que invadem o endométrio uterino; o blastocisto, então, aprofunda-se nesse endométrio. À medida que a invasão prossegue, os vasos e glândulas do endométrio podem ser corroídos por enzimas embrionárias e o sangue materno acaba jorrando nas lacunas que estão se formando. Essas lacunas fornecem a nutrição inicial e oxigênio ao embrião. No entanto, os sangues materno e embrionário não se misturam. Existe uma barreira separando-os, constituída pela parede das vilosidades.
Como se pode notar, a placenta é construída com a participação de tecidos maternos e embrionários. Ao contrário do que poderia pensar, a placenta não envolve o embrião. Essa função é exercida pelo âmnio (bolsa d’água), dentro do qual o embrião fica imerso. Esse anexo é muito desenvolvido nos mamíferos. O cório adere ao âmnio e ambos contornam a cavidade amniótica, preenchida pelo líquido amniótico.
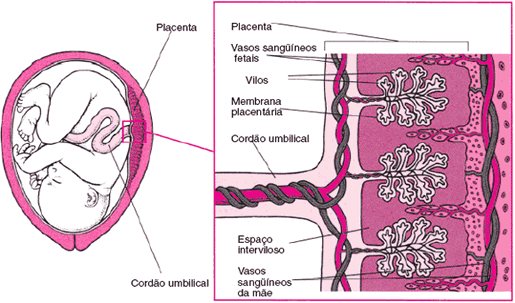
Nos mamíferos placentários, o saco vitelínico e o alantóide possuem pequeno tamanho e deixam de exercer a função desempenhada em aves e répteis. Contribuem, no entanto, para a formação do cordão umbilical, uma espécie de pedúnculo que liga a placenta ao embrião e é forrado pela membrana do âmnio, que reveste o saco vitelínico e a alantóide regredidos. No interior do cordão umbilical, duas artérias conduzem sangue do embrião para a mãe, enquanto uma veia transporta sangue em sentido contrário.
As três consequências da fecundação
A primeira consequência da fecundação é o restabelecimento da diploidia. O espermatozóide é haplóide e o óvulo também. Logo, a mistura dos lotes cromossômicos de ambos forma uma célula diplóide, a célula-ovo ou zigoto.
A segunda consequência é a determinação do sexo, uma ocorrência particularmente importante nos mamíferos.
A terceira consequência da fecundação é que ela desencadeia uma série de eventos que permitirão o desenvolvimento do zigoto em um futuro embrião. É no meio de todo esse processo, que já foi explicado anteriormente, que pode ocorrer a formação dos gêmeos.
Formação dos Gêmeos
Chama-se gêmeos dois ou mais irmãos que nascem num nascimento múltiplo, ou seja, de uma mesma gestação da mãe, podendo ser idênticos ou não. Por extensão, as crianças nascidas de partos triplos, quádruplos ou mais também são chamados de gêmeos. Apesar de não haver uma estatística precisa, estima-se que uma em cada 85 gravidezes é gemelar. Existem duas maneiras de nascerem irmãos gêmeos.
Gêmeos Bivitelinos
Os gêmeos bivitelinos são dizigóticos ou multivitelinos, ou seja, são formados a partir de dois óvulos. Nesse caso são produzidos dois ovócitos II e os dois são fecundados, formando assim, dois embriões. Quase sempre são formados em placentas diferentes e não dividem o saco amniótico.
 | Os gêmeos fraternos não se assemelham muito entre si, podem ter, ou não, o mesmo fator sanguíneo e podem ser do mesmo sexo ou não. Também são conhecidos como gêmeos diferentes. Na verdade são dois irmãos comuns que tiveram gestação coincidente. Representam 66% de todas as gestações gemelares, e neste tipo de gestação, 1/3 têm sexos diferentes, enquanto 2/3 o mesmo sexo. Um em cada um milhão de gêmeos deste tipo têm cores diferentes, mesmo sendo do mesmo pai. É possível gêmeos fraternos terem pais completamente diferentes. |
Irmãos nascidos da mesma gravidez e desenvolvidos a partir de dois óvulos que foram liberados do ovário simultaneamente e fertilizados na mesma relação sexual (regra), porém podem ser concebidos de cópulas distintas, mas daquela mesma ovulação dupla. Podem ter ou não do mesmo sexo, se diferenciam tanto fisicamente como em sua constituição genética e possuem duas placentas e duas membranas independentes e bem diferenciadas.
A frequência dos gêmeos dizigóticos varia de acordo com a origem étnica (máxima incidência na raça negra, mínima na asiática e intermediária na branca), a idade materna (máxima quando a mãe tem de 35 a 39 anos) e a genética, com uma maior incidência da linha genética materna que da paterna, ainda que os pais possam transmitir a predisposição à dupla ovulação à suas filhas. Em geral, a proporção global é de dois terços de gêmeos dizigóticos para um de monozigóticos (ou seja, os gêmeos idênticos).
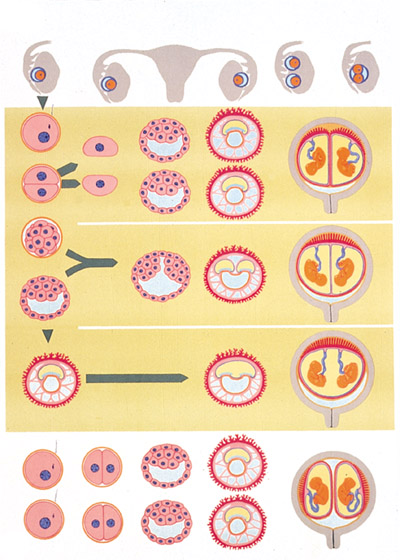
Gêmeos Idênticos
Quando um óvulo é produzido e fecundado por um só espermatozóide e se divide em duas culturas de células completas, dá origem aos gêmeos idênticos, ou monozigóticos, ou univitelinos. Sempre possuem o mesmo sexo. Os gêmeos idênticos têm o mesmo genoma, e são clones um do outro. Apenas 1/3 das gestações são de gêmeos univitelinos.
A gestação é difícil pelo fato de apenas 10% a 15% dos gêmeos idênticos terem placentas diferentes, geralmente possuem a mesma placenta.
Gêmeos xifópagos (siameses)
Os gêmeos xifópagos, ou siameses, são monozigóticos, ou seja, formados a partir do mesmo zigoto.
 | Porém, nesse caso, o disco embrionário não chega a se dividir por completo, produzindo gêmeos que estarão ligados por uma parte do corpo, ou têm uma parte do corpo comum aos dois. O embrião de gêmeos xifópagos é, então, constituído de apenas uma massa celular, sendo desenvolvido na mesma placenta, com o mesmo saco aminiótico. Estima-se que dentre 40 gestações gemelares monozigóticas, uma resulta em gêmeos interligados por não separação completa. Num outro tipo de gêmeos xifópagos (hoje sabidamente mais comum) a união acontece depois, ou seja, são gêmeos idênticos separados que se unem em alguma fase da gestação por partes semelhantes: cabeça com cabeça; abdômen com abdômen; nádegas com nádegas, etc. Quando vemos alguma notícia de gêmeos que foram "separados" por cirurgia, trata-se, quase sempre, de um caso destes. O termo "siameses" originou-se de uma famosa ocorrência registrada desse fenômeno: os gêmeos Chang e Eng, que nasceram no Sião, Tailândia, em 1811, colados pelo ombro. Eles casaram, tiveram 22 filhos e permaneceram unidos até o fim de seus dias, tendo falecido com um intervalo de 3 horas um do outro. |
www.sobiologia.com

Incrivelmente didático !
ResponderExcluir