A fotossíntese é o principal processo autotrófico e é realizada pelos seres clorofilados, representados por plantas, alguns protistas, bactérias fotossintetizantes e cianobactérias.
Na fotossíntese realizada pelos seres fotossintetizantes, com exceção das bactérias, gás carbônico (CO2) e água (H20) são usados para a síntese de carboidratos, geralmente a glicose. Nesse processo há a formação de oxigênio (O2), que é liberado para o meio. |
A fotossíntese realizada pelas bactérias fotossintetizantes difere em muitos aspectos da realizada pelos demais organismos fotossintetizantes, como veremos a seguir.
A fórmula geral da produção de glicose pela fotossíntese dos eucariotos e cianobactérias é:
6 CO2 + 12 H2O  C6H12O6 + 6 O2 + H2O
C6H12O6 + 6 O2 + H2O
Essa equação mostra que, na presença de luz e clorofila, o gás carbônico e a água são convertidos em uma hexose – neste exemplo, a glicose - havendo liberação de oxigênio.
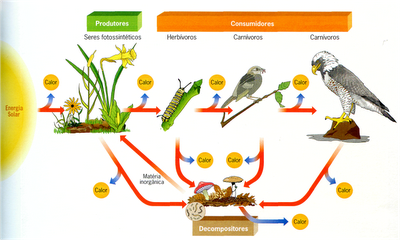
Os seres fotossintetizantes são fundamentais para a manutenção da vida em nosso planeta, pois são a base da maior parte das cadeias alimentares e produzem oxigênio, gás mantido na atmosfera em concentrações adequadas graças principalmente a atividade fotossintética.
Origem do oxigênio e fotossíntese bacteriana
O oxigênio liberado pela fotossíntese realizada pelos eucariontes e pelas cianobactérias provém da água, e não do gás carbônico, como se pensava antigamente.
O primeiro pesquisador a propor isso foi Cornelius Van Niel, na década de 1930, quando estudava bactérias fotossintetizantes. Ele verificou que as bactérias vermelhas sulfurosas (ou tiobactérias púrpuras) realizavam uma forma particular de fotossíntese em que não havia necessidade de água nem formação de oxigênio. Essas bactérias usam gás carbônico e sulfeto de hidrogênio (H2S) e produzem carboidrato e enxofre.
Van Niel escreveu, então, a fórmula geral da fotossíntese realizada por essas bactérias:
Fotossíntese bacteriana
6 CO2+ 2 H2S  CH2O + H2O + 2 S
CH2O + H2O + 2 SFoi a compreensão desse processo de fotossíntese que levou o pesquisador a propor a equação geral da fotossíntese:
6 CO2+ 2 H2A  CH2O + H2O + 2 A
CH2O + H2O + 2 A Essa equação mostra que H2A pode ser a água (H2O) ou o sulfeto de hidrogênio (H2S) e evidencia que, se for água ela é a fonte de oxigênio na fotossíntese.
Essa interpretação foi confirmada posteriormente, na década de 1940, por experimentos em que pesquisadores forneciam às plantas água cujo oxigênio era de massa 18 (O18, isótopo pesado do oxigênio) em vez de 16 (O16), como o oxigênio da água comum. Eles verificaram que o oxigênio liberado pela fotossíntese era o O18, corroborando a interpretação de Van Niel.
Ficou comprovado, então, que o oxigênio liberado durante a fotossíntese dos eucariontes e das cianobactérias provém da água e não do gás carbônico.
Onde ocorre a fotossíntese?
Nos organismos mais simples, como as cianobactérias, a fotossíntese ocorre no hialoplasma, que é onde se encontram diversas moléculas de clorofila, associadas a uma rede interna de membranas, que são extensões da membrana plasmática. Recorde que as cianobactérias são procariontes e não possuem organelas dotadas de membranas. Por outro lado, nos organismos eucariontes a fotossíntese ocorre totalmente no interior do cloroplasto.
Os Cloroplastos
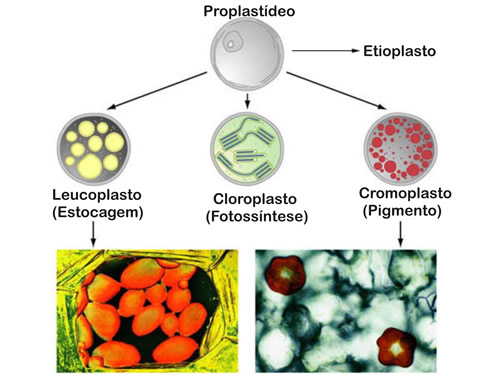
Os plastos ou plastídeos é um grupo de organelas específicas de células vegetais, que possuem características semelhantes com as mitocôndrias como: membrana dupla, DNA próprio e origem endosimbionte.
Os plastos desenvolvem-se a partir de proplastídeos, que são organelas pequenas presentes nas células imaturas dos meristemas vegetais e desenvolvem-se de acordo com as necessidades da célula, surgindo diferentes tipos de plastos como: os cromoplastos (que contêm pigmentos), os leucoplastos (sem pigmento), etioplastos (que se desenvolvem na ausência de luz), amiloplastos (que acumulam amido como substância de reserva), proteoplastos (que armazenam proteína) e os oleoplastos (acumulam lipídeos).
Os cloroplastos são um tipo de cromoplastos que contém pigmento chamado clorofila, que são capazes de absorver a energia eletromagnética da luz solar e a convertem em energia química por um processo chamado fotossíntese.
As células vegetais e as algas verdes possuem um grande número de cloroplastos, de forma esférica ou ovóide, variando de tamanho de acordo com o tipo celular, e são bem maiores que as mitocôndrias.
Origem evolutiva dos cloroplastos
Acredita-se que os cloroplastos tenham se originado de organismos procariontes fotossintéticos (algas azuis), que se instalaram em células primitivas eucariontes aeróbicas por endossimbiose. Essa simbiose há cerca de 1,2 bilhões de anos, teria dado origem às algas vermelhas, depois as algas pardas e verdes e aos vegetais superiores.
Durante o processo evolutivo, as bactérias precursoras dos cloroplastos transferiram parte de seu material genético para o DNA da célula hospedeira, assim passaram a depender do genoma da célula hospedeira para a produção de muitas de suas proteínas.
Esta origem é semelhante ao da mitocôndria, mas existem diferenças como o tamanho das organelas, o cloroplasto é bem maior que a mitocôndria, e a fonte de energia é diferente, o cloroplasto usa energia luminosa enquanto a mitocôndria usa energia química.
Composição química dos cloroplastos Os cloroplastos são as organelas mais evidentes das células vegetais. Ela é composta por 50% de proteínas, 35% de lipídeos, 5% de clorofila, água e carotenóides. Parte das proteínas são sintetizadas pelo núcleo da célula, mas os lipídeos são sintetizados dentro da própria organela.
O número de cloroplastos é regulado pela célula. Existem células que contém apenas um cloroplasto, mais a maioria das células que realizam fotossíntese contém cerca de 40 a 200 cloroplastos, que se movimentam em função da intensidade de luz e da corrente citoplasmática.
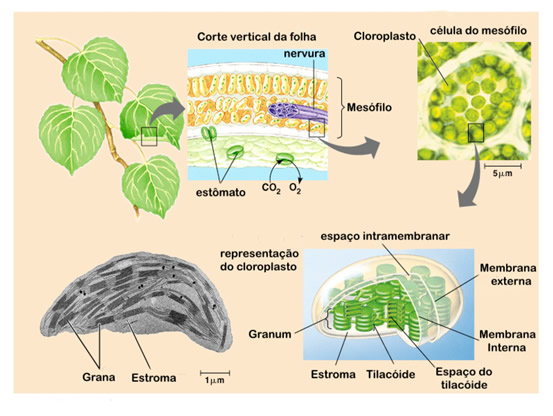
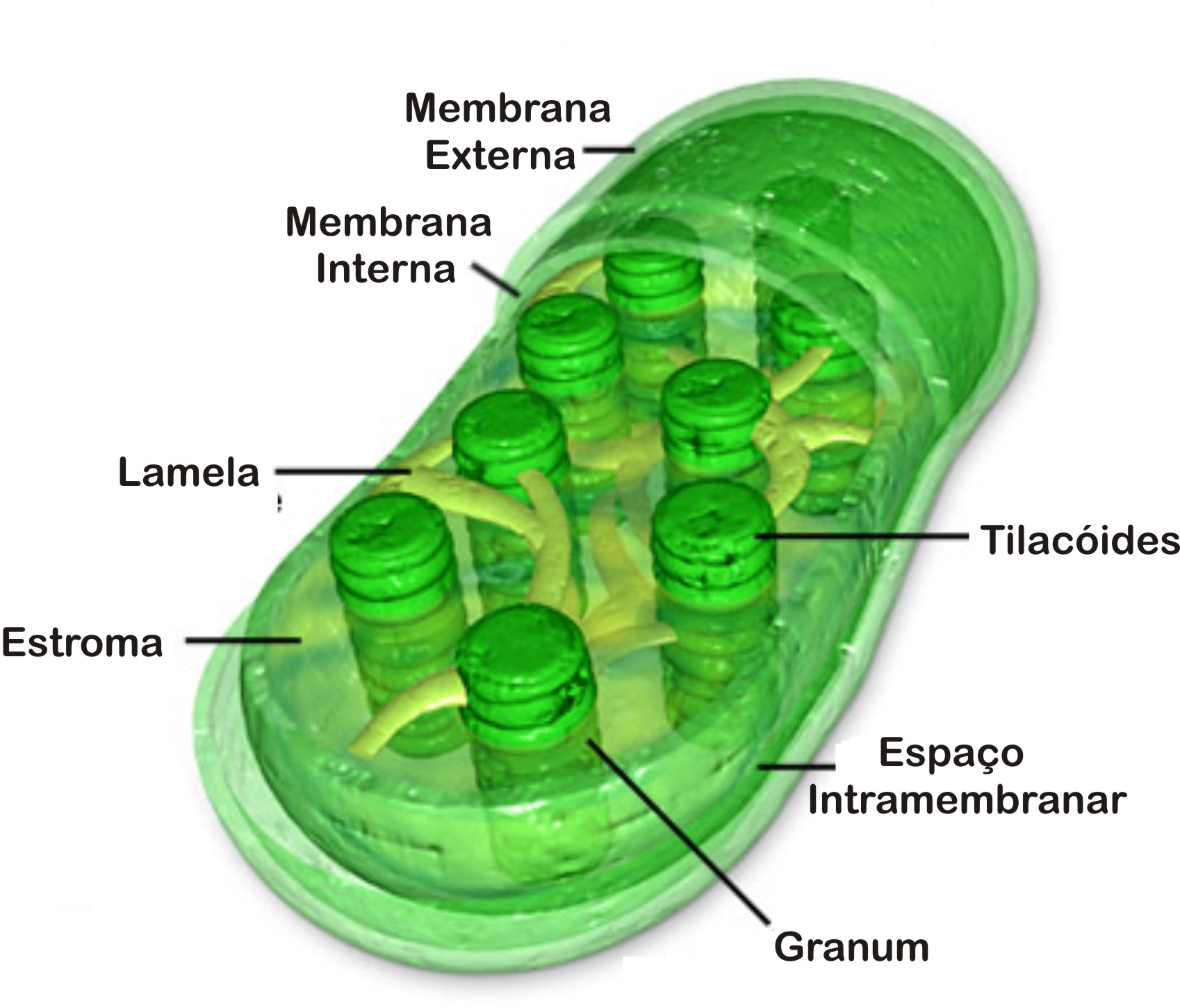
Semelhantes às mitocôndrias, os cloroplastos são envoltos por duas membranas, uma externa altamente permeável, e uma interna que necessita de proteínas específicas para o transporte de metabólicos, e um espaço intermembrana. No interior da organela existe uma matriz amorfa chamada estroma que contém várias enzimas, grãos de amido, ribossomos e DNA. | | 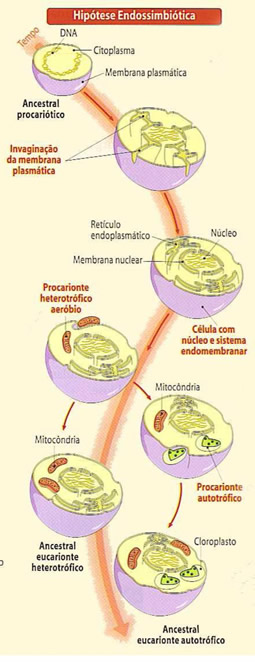 |
No entanto, a membrana interna do cloroplasto não é dobrada em cristas e não contém uma cadeia transportadora de elétrons. Mergulhado no estroma, existe um sistema de membrana (bicamada) que forma um conjunto de sacos achatados em forma de discos chamados de membrana tilacóide (do grego thylakos, saco).
O conjunto de discos empilhados recebe o nome de granum. O lúmen da membrana tilacóide é chamado de espaço tilacóide. Na membrana exposta ao estroma se localizam as clorofilas que participam da fotossíntese.
Os pigmentos ligados a diferentes proteínas e lipídeos nas membranas dos tilacóides granares e estromáticos formam sistemas complexos de proteínas-clorofila denominados fotossistemas. Há dois tipos de fotossistemas:
Fotossistema I: localizado na região da membrana voltada para o estroma, são as menores partículas intramembranosas.
Fotossistema II: localizado em tilacóides granares, formado por partículas maiores.
Sistema genético dos cloroplastos
O genoma plastidial consiste em uma pequena molécula de DNA circular, com características muito semelhantes com das mitocôndrias e das bactérias.
O DNA dos plastos ocorre em maior quantidade e é mais complexo do que da mitocôndria. Existem 30 a 200 cópias de DNA por organela contendo aproximadamente 120 genes.
O sequenciamento genético dos cloroplastos de várias plantas levou a identificação de muitos desses genes. Eles transcrevem todos os RNAs ribossômicos que compõem os plastoribossomos e 30 tipos diferentes de RNA transportadores.
Esse genoma codifica ainda 20 proteínas ribossômicas, 30 proteínas que funcionam na fotossíntese e algumas subunidades de RNA polimerase (proteínas envolvidas na expressão gênica).
Mas mesmo sintetizando suas próprias proteínas, cerca de 90% das proteínas dos cloroplastos são codificadas pelos genes nucleares que são importadas do citosol para a organela.
Luz - Componente indispensável da fotossíntese
A luz que banha a Terra é componente do amplo espectro de radiações eletromagnéticas provenientes do Sol, e que se propagam como ondas. O modo como essas ondas se propagam depende da energia: quanto mais energia uma onda tiver, menor será seu comprimento.
Dentro do amplo espectro de radiações eletromagnéticas, apenas uma pequena parte é visível aos nossos olhos – são as radiações cujos comprimentos de onda vão de 380 a 760 nanômetros. Essas estreitas faixas de comprimento de onda da luz visível corresponde às diferentes cores que são observadas quando se faz passar a luz por um prisma, o que provoca a dispersão (separação) dessas diferentes radiações.
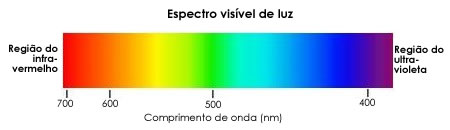
Outra característica importante da luz é a sua natureza corpuscular, ou seja, a luz é característica por incidir na forma de corpúsculos, conhecidos como fótons. Os fótons são considerados “pacotes” de energia associados a cada comprimento de onda partícula. Luz de pequeno comprimento de onda, como a luz violeta, possui fótons altamente energéticos. Luz de grande comprimento de onda, como a vermelha e a laranja, possuem fótons pouco energéticos. Assim, cada radiação luminosa, cada comprimento de onda luminosa, é portadora de uma certa energia. E o fato notável é que as plantas aproveitam essa energia para a produção de matéria orgânica na fotossíntese.
Como isso pode ser provado?
Se um feixe de luz branca passar por um prisma se decompondo em diversas cores, e em cada cor colocarmos um vidro cheio de água, lacrado, e com uma plantinha dentro, veremos com o decorrer do tempo que as plantas submetidas às radiações vermelha e azul mostram o mais alto grau de atividade fotossintética. Isso pode ser percebido pelo tamanho da bolha de oxigênio que se formou em cada vidro.
Pigmentos fotossintetisantes
O termo "pigmento" significa substância colorida. A cor do pigmento fotossintetizante depende das faixas do espectro da luz visível que ele absorve ou reflete.
A clorofila, que dá a cor verde característica da maioria dos vegetais, absorve muito bem a luz nas faixas do vermelho e do violeta, refletindo a luz verde.
Como a luz refletida é a que atinge os nossos olhos, essa é a cor que vemos, ao olharmos para uma folha. O perfil de absorção de luz de uma substância é o seu espectro de absorção.
Todas as células fotossintetizantes, exceto as bacterianas, contêm 2 tipos de clorofila, e um deles sempre é a clorofila a. O segundo tipo de clorofila geralmente é a clorofila b (nos vegetais superiores) ou a clorofila c (em muitas algas). Esses diversos tipos de clorofila diferem quanto à faixa do espectro da luz visível na qual cada uma delas capta luz com mais eficiência.
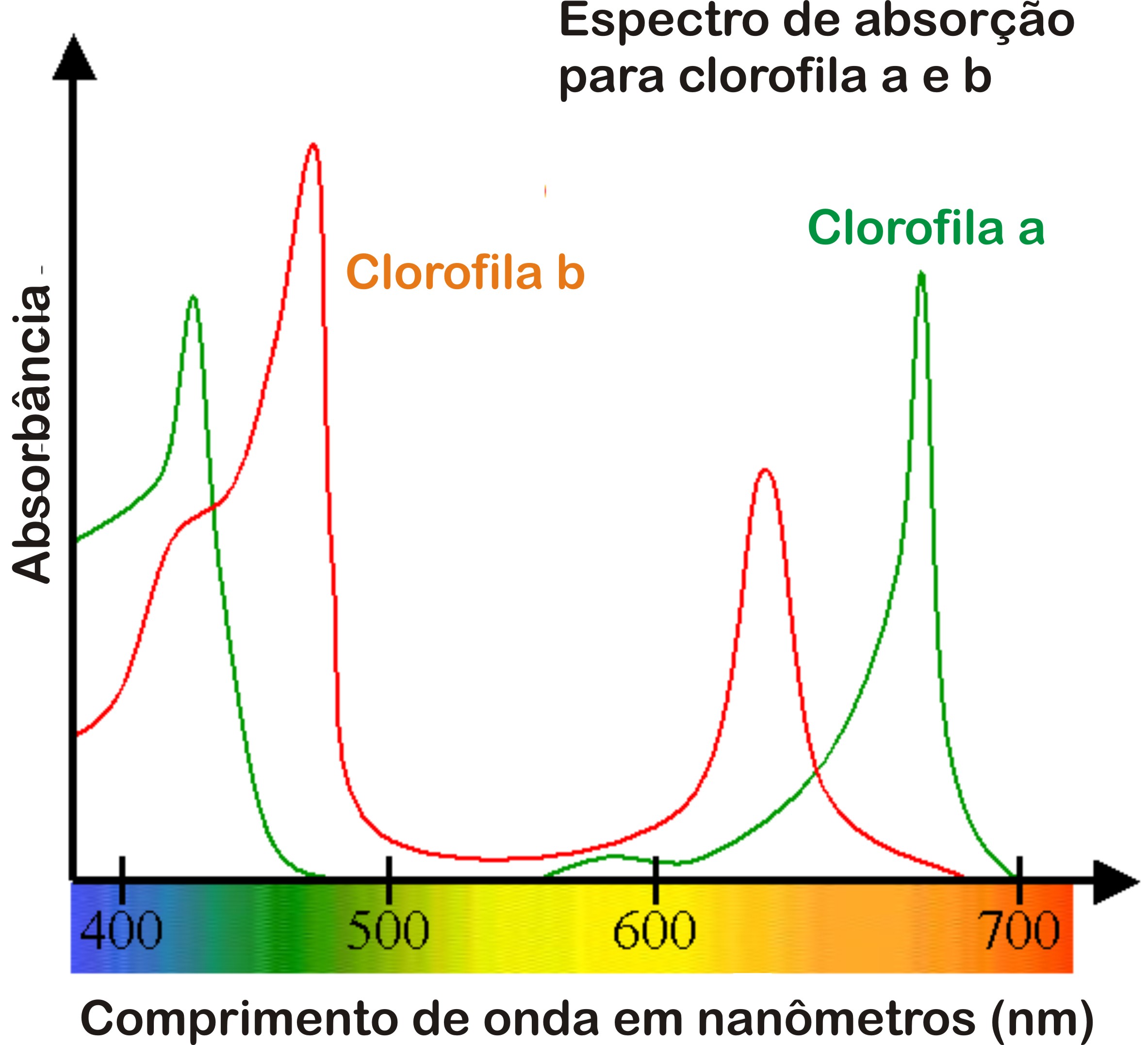
As clorofilas a e b possuem espectros de absorção de luz ligeiramente diferentes, como mostra o gráfico a seguir:
Podemos verificar, analisando o gráfico, que ambas as clorofilas possuem dois picos de absorção: um mais elevado, na faixa do violeta, e um outro menor, na faixa do vermelho.
Os carotenóides são pigmentos acessórios. Eles absorvem luz em faixas um pouco diferentes das faixas das clorofilas. A presença desses pigmentos acessórios faz com que muitas folhas tenham cores diferentes do verde. Embora tenham clorofila, a presença desses outros pigmentos em grandes quantidades mascara a sua presença e deixa as folhas com outras cores (arroxeadas, alaranjadas, amarelas, etc.).
Muitas folhas mudam de cor, no inverno, pela diminuição na quantidade de clorofila. Como a quantidade dos outros pigmentos não se altera tão significativamente, as suas cores passam a ser vistas, tornando as folhas geralmente amareladas.
O papel da luz na fotossíntese
A estrutura atômica de determinadas substâncias é tal que as tornam capazes de absorver a luz. Quando a luz incide em um átomo capaz de absorvê-la, alguns elétrons são ativados e elevados a um nível energético superior. O átomo entra em um "estado ativado", rico em energia e muito instável. Quando os elétrons excitados voltam aos seus orbitais normais, o átomo volta ao seu estado-base. Esse retorno é acompanhado pela liberação de energia, como calor ou como luz. A luz emitida dessa forma é chamada fluorescência.
Nos cloroplastos, as moléculas de clorofila possuem essa característica. Entretanto, o seu elétron excitado não devolve a energia captada através da fluorescência, mas a transfere para outras substâncias. Há, portanto, transformação da energia luminosa captada em energia química.
As etapas da fotossíntese
A fotossíntese ocorre em duas grandes etapas, que envolvem várias reações químicas: a primeira é a fase clara (também chamada de fotoquímica) e a segunda é a fase escura (também conhecida como fase química).
Em linhas gerais, os eventos principais da fotossíntese são a absorção da energia da luz pela clorofila; a redução de uma aceptor de elétrons chamado NADP, que passa a NADPH2; a formação de ATP e a síntese de glicose.
A fase escura da fotossíntese não precisa ocorrer no escuro. O que o nome quer indicar é que ela ocorre mesmo na ausência de luz – ela só precisa de ATP e NADH2 para ocorrer.
Fase clara ou fotoquímica: Quebra da água e liberação de oxigênio
Esta fase ocorre na membrana dos tilacóides e dela participam um complexo de pigmentos existente nos grana, aceptores de elétrons, moléculas de água e a luz. Como resultado desta fase temos a produção de oxigênio, ATP (a partir de ADP + Pi) e também a formação de uma substância chamada NADPH2;. Tanto o ATP quanto o NADPH2; serão utilizadas na fase escura.
Na fase clara, a luz penetra nos cloroplastos e atinge o complexo de pigmentos, ao mesmo tempo em que provoca alterações nas moléculas de água. De que maneira essa ação da luz resulta em produtos que podem ser utilizadas na segunda fase da fotossíntese?
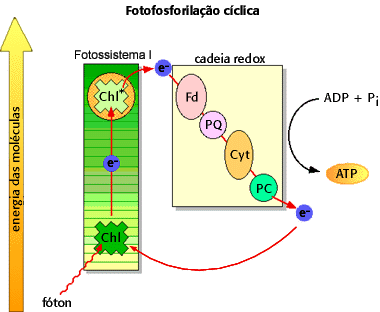
Um dos acontecimentos marcantes da fase clara são as chamadas fotofosforilações cíclica e acíclica.
Na fotofosforilação cíclica, ao ser atingida pela luz do Sol, a molécula de clorofila libera elétrons. Esses elétrons são recolhidos por determinadas moléculas orgânicas chamadas aceptores de elétrons, que os enviam a uma cadeia de citocromos (substâncias associadas ao sistema fotossintetizante e que são assim chamadas por possuírem cor). Daí, os elétrons retornam à clorofila.
Você poderá perguntar: qual a vantagem desse ciclo de transporte de elétrons?
A resposta é que ao efetuar o retorno para a molécula de clorofila, a partir dos citocromos, os elétrons liberam energia, pois retornam aos seus níveis energéticos originais. E essa energia é aproveitada para a síntese de moléculas de ATP, que serão utilizadas na fase escura da fotossíntese.
Perceba que o caminho executado pelos elétrons é cíclico. Por esse motivo, costuma-se denominar essa via de fotofosforilação cíclica, devido à ocorrência de síntese de inúmeras moléculas de ATP em um processo cíclico, com a participação da luz e de moléculas de clorofila.
Ao mesmo tempo que isso ocorre, moléculas de água – ao serem atingidas pela luz do Sol – são “quebradas” (usa-se o termo “fotólise da água” para designar a quebra das moléculas de água) e liberam prótons (H+), elétrons (e-) e moléculas de oxigênio. Os prótons são captados por moléculas de NADP, que se convertem em NADPH2; moléculas de oxigênio são liberados para o meio; e os elétrons voltam para a clorofila, repondo aqueles que ela perdeu no início do processo.
A Etapa Fotoquímica da Fotossíntese
Também é chamada "fase clara" da fotossíntese, uma vez que a sua ocorrência é totalmente dependente da luz. Como se trata de uma etapa que conta com a participação das moléculas de clorofila, acontece no interior dos tilacóides, em cujas faces internas de suas membranas as moléculas desse pigmento fotossintetizante estão "ancoradas".
Nessa etapa, a clorofila, ao ser iluminada, perde elétrons, o que origina "vazios" na molécula. O destino dos elétrons perdidos e a reocupação desses vazios podem obedecer a 2 mecanismos distintos, chamados fotofosforilação cíclica e fotofosforilação acíclica.
A - Fotofosforilação cíclica
No chamado fotossistema I, predomina a clorofila a. Essa, ao ser iluminada, perde um par de elétrons excitados (ricos em energia). Estabelece-se, na molécula da clorofila, um "vazio" de elétrons. O par de elétrons é recolhido por uma série de citocromos, substâncias que aceitam elétrons adicionais, tornando-se instáveis e transferindo esses elétrons para outras moléculas.
À medida que passam pela cadeia de citocromos, os elétrons vão gradativamente perdendo energia, que é empregada na fosforilação (produção de ATP pela união de mais um grupo de fosfato a uma molécula de ADP). Como essa fosforilação é possível graças à energia luminosa, captada pelos elétrons da clorofila, é chamada fotofosforilação.
Após a passagem pela cadeia de citocromos, os elétrons retornam à molécula da clorofila, ocupando o "vazio" que haviam deixado. Como os elétrons retornam para a clorofila, o processo é
cíclico.
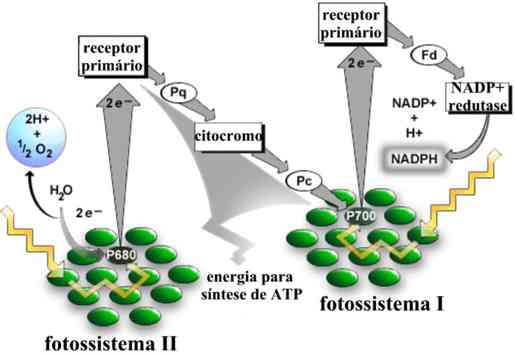
B - Fotofosforilação acíclica
Esse mecanismo emprega dois sistemas fotossintetizantes: o fotossistema I e o fotossistema II. No fotossistema I, predomina a clorofila a, enquanto no fotossistema II, predomina a clorofila b.
A clorofila a, iluminada, perde um par de elétrons ativados, recolhidos por um aceptor especial, a ferridoxina. Ao mesmo tempo, a clorofila b, excitada pela luz, perde um par de elétrons que, depois de atravessarem uma cadeia de citrocromos, ocupa o "vazio" deixado na molécula da clorofila a. Durante a passagem desses elétrons pela cadeia de citocromos, há liberação de energia e produção de ATP (fosforilação). Como o "vazio de elétrons" da clorofila a não é preenchido pelos mesmos elétrons que saíram dessa molécula, o mecanismo é chamado fotofosforilação acíclica.
No interior dos cloroplastos, a água é decomposta na presença da luz. Essa reação é a fotólise da água. (ou reação de Hill).
Dos produtos da fotólise da água, os elétrons vão ocupar os "vazios" deixados pela perda de elétrons pela clorofila b. Os prótons H+, juntamente com os elétrons perdidos pela clorofila a, irão transformar o NADP (nicotinamida-adenina-dinucleotídeo fosfato) em NADPH. Ao mesmo tempo, oxigênio é liberado. Esse é um aspecto importante da fotossíntese: todo o oxigênio gerado no processo provém da fotólise da água.
Os seres fotossintetizantes utilizam a água como fonte de átomos de hidrogênio para a redução do NADP. Esses átomos de hidrogênio são posteriormente empregados na redução do CO2 até carboidrato. A equação geral do processo é a seguinte:
O valor n corresponde, geralmente, a seis, o que leva à formação de glicose (C6H12O6). Entretanto, como todo oxigênio liberado vem da água, a equação deve ser corrigida para:
Dessa forma, pode-se explicar a origem de uma quantidade 2n de átomos de oxigênio a partir de uma quantidade de 2n moléculas de água (H2O).
Fase escura ou química: Produção de Glicose
Nessa fase, a energia contida nos ATP e os hidrogênios dos NADPH2, serão utilizados para a construção de moléculas de glicose. A síntese de glicose ocorre durante um complexo ciclo de reações (chamado ciclo das pentoses ou ciclo de Calvin-Benson), do qual participam vários compostos simples.
Durante o ciclo, moléculas de CO2 unem-se umas as outras formando cadeias carbônicas que levam à produção de glicose. A energia necessária para o estabelecimento das ligações químicas ricas em energia é proveniente do ATP e os hidrogênio que promoverão a redução dos CO2 são fornecidos pelos NADPH2.
Veja com mais detalhes o ciclo de Calvin
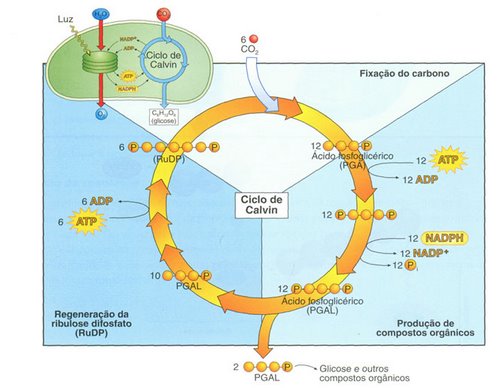
O Ciclo de Calvin
O ciclo começa com a reação de uma molécula de CO2 com um açúcar de cinco carbonos conhecido como ribulose difosfato catalisada pela enzima rubisco (ribulose bifosfato carboxilase/oxigenase, RuBP), uma das mais abundantes proteínas presentes no reino vegetal.
Forma-se, então, um composto instável de seis carbonos, que logo se quebra em duas moléculas de três carbonos (2 moléculas de ácido 3-fosfoglicérico ou 3-fosfoglicerato, conhecidas como PGA). O ciclo prossegue até que no final, é produzida uma molécula de glicose e é regenerada a molécula de ribulose difosfato.
Note, porém, que para o ciclo ter sentido lógico, é preciso admitir a reação de seis moléculas de CO2 com seis moléculas de ribulose difosfato, resultando em uma molécula de glicose e a regeneração de outras seis moléculas de ribulose difosfato.
A redução do CO2 é feita a partir do fornecimento de hidrogênios pelo NADH2 e a energia é fornecida pelo ATP. Lembre-se que essas duas substâncias foram produzidas na fase clara.
O esquema apresentado é uma simplificação do ciclo de Clavin: na verdade, as reações desse ciclo se parecem com as que ocorrem na glicólise, só que em sentido inverso.
É correto admitir, também, que o ciclo origina unidades do tipo CH2O, que poderão ser canalizadas para a síntese de glicose, sacarose, amido e, inclusive, aminoácidos, ácidos graxos e glicerol.
Fatores que influenciam a fotossíntese
A intensidade com a qual uma célula executa a fotossíntese pode ser avaliada pela quantidade de oxigênio que ela libera para o ambiente, ou pela quantidade de CO2 que ela consome.
Quando se mede a taxa de fotossíntese de uma planta, percebe-se que essa taxa pode aumentar ou diminuir, em função de certos parâmetros. Esses parâmetros são conhecidos como fatores limitantes da fotossíntese. A fotossíntese tem alguns fatores limitantes, alguns intrínsecos e outros extrínsecos.
Fatores limitantes intrínsecos
Disponibilidade de pigmentos fotossintetizantes
Como a clorofila é a responsável principal pela captação da energia limunosa, a sua falta restringe a capacidade de captação da energia e a possibilidade de produzir matéria orgânica.
Disponibilidade de enzimas e de cofatores
Todas as reações fotossintéticas envolvem a participação de enzimas e de co-fatores, como os aceptores de elétrons e os citocromos. A sua quantidade deve ser ideal, para que a fotossíntese aconteça com a sua intensidade máxima.
Fatores limitantes extrínsecos
A concentração de CO2
 | | O CO2 (gás carbônico ou dióxido de carbono) é o substrato empregado na etapa química como fonte do carbono que é incorporado em moléculas orgânicas. As plantas contam, naturalmente, com duas fontes principais de CO2: o gás proveniente da atmosfera, que penetra nas folhas através de pequenas aberturas chamadas estômatos, e o gás liberado na respiração celular.
Sem o CO2, a intensidade da fotossíntese é nula. Aumentando-se a concentração de CO2 a intensidade do processo também se eleva. Entretanto, essa elevação não é constante e ilimitada. Quando todo o sistema enzimático envolvido na captação do carbono estiver saturado, novos aumentos na concentração de CO2 não serão acompanhados por elevação na taxa fotossintética. |
A Temperatura
 | |
Na etapa química, todas as reações são catalisadas por enzimas, e essas têm a sua atividade influenciada pela temperatura.
De modo geral, a elevação de 10 °C na temperatura duplica a velocidade das reações químicas.
Entretanto, a partir de temperaturas próximas a 40 °C, começa a ocorrer desnaturação enzimática, e a velocidade das reações tende a diminuir.
Portanto, existe uma temperatura ótima na qual a atividade fotossintetizante é máxima, que não é a mesma para todos os vegetais. |
O comprimento de onda
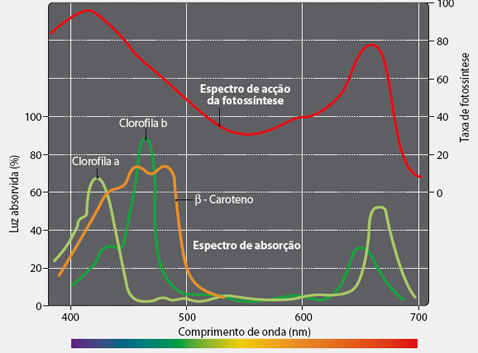
| | A assimilação da luz pelas clorofilas a e b, principalmente, e secundariamente pelos pigmentos acessórios, como os carotenóides, determina o espectro de ação da fotossíntese.
Nota-se a excelente atividade fotossintética nas faixas do espectro correspondentes à luz violeta/azul e à luz vermelha, e à pouca atividade na faixa do verde.
Para que uma planta verde execute a fotossíntese com boa intensidade, não se deve iluminá-la com luz verde, uma vez que essa luz é quase completamente refletida pelas folhas. |
Intensidade luminosa
 | | Quando uma planta é colocada em completa obscuridade, ela não realiza fotossíntese. Aumentando-se a intensidade luminosa, a taxa da fotossíntese também aumenta. Todavia, a partir de um certo ponto, novos aumentos na intensidade de iluminação não são acompanhados por elevação na taxa da fotossíntese. A intensidade luminosa deixa de ser um fator limitante da fotossíntese quando todos os sistemas de pigmentos já estiverem sendo excitados e a planta não tem como captar essa quantidade adicional de luz. Atingiu-se o ponto de saturação luminosa.
Aumentando-se ainda mais a intensidade de exposição à luz, chega-se a um ponto a partir do qual a atividade fotossintética passa a ser inibida. Trata-se do ponto de inibição da fotossíntese pelo excesso de luz. |