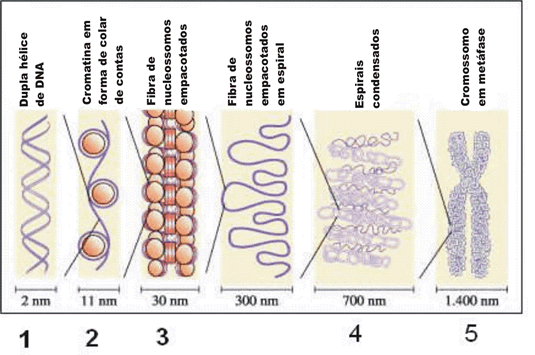
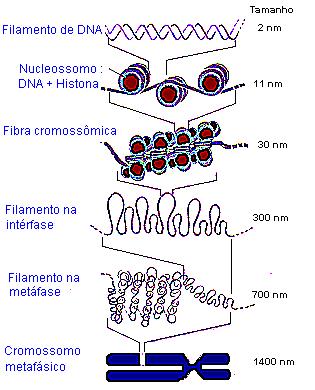
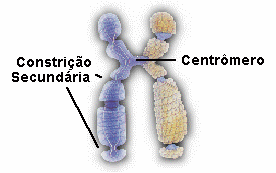
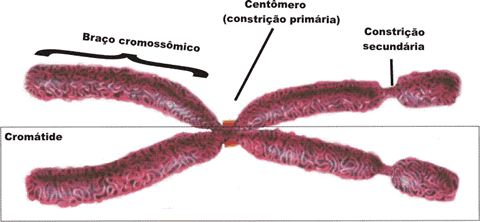
OS SERES VIVOS
A distribuição da vida na biosfera
A distribuição da vida na biosfera
A fina camada de solo, água e ar que abriga a vida em nosso planeta é chamada biosfera.
Na biosfera encontramos ambientes muito diferentes, que vão desde os oceanos com profundidades que atingem nove mil metros até as montanhas com mais de oito mil metros de altitude. Em todos esses locais existem formas de vida.
É claro que cada tipo de ambiente da biosfera apresenta condições abióticas específicas, propiciando a vida de comunidades diferentes e formando, assim, ecossistemas diferenciados.
A vida nos mares
A salinidade, a temperatura e a luminosidade são fatores importantes para a distribuição da vida no ambiente marinho.
Nas águas dos mares, que cobrem mais de 70% da superfície do globo terrestre, encontramos várias substâncias químicas dissolvidas. A principal delas é o cloreto de sódio ou sal comum.
O conteúdo de sais dissolvidos na água do mar determina sua salinidade, que pode variar muito, dependendo da quantidade de água doce proveniente dos rios que ali desembocam e do grau de evaporação da água.
As radiações solares que chegam até o planeta produzem efeitos de luz e calor sobre os mares. Esses efeitos variam com a profundidade: quanto mais profundas forem as regiões do mar, menos luz e calor elas recebem. Por causa disso, surgem regiões muito diferentes, que tornam possível a existência de uma grande variedade de seres vivos. Podemos assim observar três regiões distintas: eufótica, disfótica e afótica.
Zona eufótica - Região de grande luminosidade, que vai até aproximadamente oitenta metros de profundidade. Aí a luz penetra com grande intensidade, possibilitando um ambiente favorável à vida de organismos fotossintetizantes, como as algas, e muitos animais que se alimentam delas.
Zona disfótica - Região em que a luz apresenta dificuldade de penetrar, tornando-se difusa. Esta região vai até cerca de duzentos metros de profundidade e também abriga organismos fotossintetizantes, embora em proporção menor que a da zona eufótica.
Zona afótica - Região totalmente escura, que vai além dos duzentos metros de profundidade. Aí não é possível a existência de animais herbívoros.
As comunidades dos seres vivos marinhos
Dependendo do modo como se locomovem, os seres vivos marinhos são classificados em três grupos distintos: plâncton, nécton e bentos.
Flâncton - O plâncton representa o conjunto de todos os seres vivos flutuantes que são levados pelas correntezas marinhas. Eles não possuem órgãos de locomoção e, quando os têm, são rudimentares. Existem duas categorias de seres planctônicos: o fitoplâncton e o zooplâncton.
- Fitoplâncton – É constituído pelos produtores, ou seja, os seres autotróficos, que desempenham um grande papel nas cadeias alimentares marinhas. As algas são os principais representantes dessa categoria.
- Zooplâncton – É constituído por organismos heterotróficos, como microcrustáceos, larvas de peixes, protozoários, insetos, pequenos anelídeos e até caravelas.
Nécton – Compreende o conjunto dos seres que nadam livremente, deslocando-se por atividade própria, vencendo a correnteza. O nécton abrange peixes (tubarões, robalos, tainhas, sardinhas, etc.), répteis, como a tartaruga, e inúmeros mamíferos (baleia, focas, golfinhos, etc.), entre outros animais.
Bentos – São o conjunto de seres que vivem fixos ou se arrastam no fundo do mar. Enfim, são os seres que pouco se afastam do fundo. Muitas algas, esponjas, ouriços-do-mar, estrelas-do-amor são exemplos de representantes de seres bentônicos.
A grande cadeia alimentar marinha
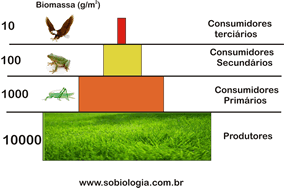
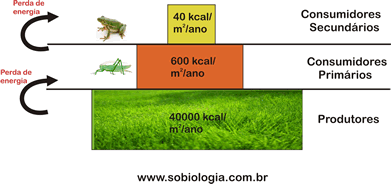
Toda a vida no mar depende da atividade fotossintetizante dos seres autróficos, principalmente das do fitoplâncton. As algas, portanto, representam o primeiro nível trófico de praticamente todas as cadeias alimentares marinhas.
Na região iluminada vivem animais herbívoros e carnívoros, além de alguns detritívoros, isto é, que se nutrem de detritos orgânicos formados por restos de organismos mortos. As cadeias alimentares marinhas são muito diversificadas e podem começar com seres autotróficos muito pequenos, como as algas unicelulares, e terminar com animais de grande porte, como tubarões e baleias.
Na região escura não existem seres fotossintetizantes e animais herbívoros. Os peixes abissais, por exemplo, que vivem em grandes profundidades, são detritívoros ou carnívoros e têm adaptações especiais para a vida nesse ambiente.
A vida nas águas continentais
As águas existentes nos continentes, rios, lagos e pântanos são denominadas águas continentais. Elas representam menos que 3% da massa de água existente no planeta. Sua temperatura varia mais que a da água dos mares e sua composição depende do tipo de solo que as suporta. O teor de salinidade é baixo e a penetração de luz é pequena.
Rios, lagos e pântanos diferem entre si pela movimentação das águas. Nos rios, as águas estão em constante mistura por causa das correntezas; nos lagos e pântanos, elas estão praticamente paradas. Considerando esse conjunto, pode-se afirmar que esses ecossistemas abrigam uma considerável diversidade de vida, que inclui algas e outros tipos de plantas, peixes, anfíbios, répteis, moluscos, anelídeos e outros animais.
Em geral, os ecossistemas de água parada produzem, através dos organismos fotossintetizantes que abrigam, o alimento necessário para a sua manutenção. Os ecossistemas de água corrente, por sua vez, são relativamente pobres em fitoplâncton. Assim, uma parte da matéria orgânica necessária para a sobrevivência dos animais que neles existem é importada dos ecossistemas terrestres vizinhos.
Infelizmente, os ecossistemas aquáticos são vítimas constantes de inúmeros resíduos originados pelos diversos tipos de atividade humana. Recebem diariamente toneladas de lixo e de esgoto doméstico, agrotóxicos, metais pesados, detergentes, etc. Alguns dos nossos rios, como o Tietê, estão enquadrados entre os mais dramáticos exemplos de poluição aquática no planeta.
Mangues, berçários da natureza
Os mangues – ambiente típico dos litorais tropicais são verdadeiros pontos de ligação entre o ambiente marinho, o de água doce e o terrestre.
Situam-se na região denominada entremarés, que se localiza entre o ponto mais alto da maré alta e o ponto mais baixo da maré baixa. Nessa região ocorre uma intensa deposição de detritos e sedimentos que, misturados à água doce e salgada, juntam-se à argila, formando um solo lamacento.
O solo dos mangues é pantanoso e movediço e possui pouco oxigênio e alta salinidade; abriga, então, plantas halófitas (que se desenvolvem em terrenos salgados), como o mangue-vermelho, com raízes-escoras ou suportes, que promovem uma eficiente fixação da planta no solo. Outra planta típica dos mangues é a Avicenia tomentosa, planta arbórea que possui raízes respiratórias; partindo da raiz principal da planta e crescendo para cima, essas raízes emergem do solo e coletam o oxigênio atmosférico, compensando o baixo teor de oxigênio do solo.
A fauna dos manguezais inclui animais como peixes, crustáceos, moluscos e aves diversas, entre outros. Você já sentiu o cheiro que se desprende de um mangue, aquele cheiro desagradável de ovo podre? Ele é produzido pelo gás sulfídrico, que resulta da ação de bactérias na decomposição de restos de animais e vegetais mortos, trazidos pelos rios e pelo mar. Essa ação decompositora das bactérias torna os mangues ricos em nutrientes. Ali muitas espécies de peixes, crustáceos e aves aquáticas vêm abrigar-se e reproduzir-se. Além da alimentação em abundância, os filhotes encontram proteção contra predadores entre as raízes das plantas e nas águas escuras. Por isso os mangues são considerados berçários da natureza.
A vida nas florestas
As florestas constituem formações vegetais em que se encontra uma quantidade enorme de nutrientes e uma diversidade muito grande de formas de vida.
Quando estudamos as florestas, um dos fatores importantes a considerar é sua estratificação, ou seja, a distribuição vertical dos vegetais. Na parte mais baixa, junto ao solo, temos a vegetação herbácea, seguindo-se a arbustiva e, finalmente, a arbórea.
Como as florestas ocupam lugares de temperaturas e climas muito diferentes, que vão desde regiões quentes e úmidas até regiões frias e secas, a sua vegetação difere bastante de um lugar para o outro. Assim, podemos distinguir três tipos básicos de floresta:
· Floresta de coníferas;
· Floresta decídua temperada;
· Floresta úmida tropical.
Floresta de coníferas do hemisfério norte
A floresta de coníferas do hemisfério norte, também denominada taiga, estende-se pelo norte da Europa, Alasca, Canadá e Sibéria.
Sua vegetação é constituída predominantemente de gimnospermas do grupo das coníferas, como os pinheiros. As coníferas não perdem as folhas durante o inverno. As folhas revestidas com cera e de pequena superfície (finas e compridas) contribuem com a redução de água por transpiração e constituem uma adaptação dessas plantas na defesa contra a insignificante absorção de água no inverno rigoroso, já que, nessas condições, a maior parte da água fica congelada no solo. Por isso, as florestas de coníferas estão sempre verdes.
A vegetação rasteira é pouco desenvolvida e formada por algumas ervas, samambaias e musgos. Isto se deve à pouca quantidade de luz que chega ao solo, pois a copa das árvores forma uma cobertura que filtra os raios luminosos do sol. Alguns tipos de fungo desenvolvem-se sobre as folhas e os ramos que caem ao solo.
A fauna é constituída de alces, ursos pardos, lobos, martas, linces, esquilos, raposas e diversas aves, entre outros animais.
As aves que habitam essas florestas geralmente alimentam-se de sementes das coníferas. É o caso do cruza-bicho, que graças ao seu bico curvo e de pontas cruzadas, consegue cortar as pinhas e abrir as sementes.
Floresta decídua temperada
Esta floresta tem uma característica marcante: as várias espécies de árvores que a constituem perdem suas folhas no final do outono, o que impede que elas se desidratem. Por isso, recebem o nome de florestas decididas.
O termo temperado aplicado a esta floresta deve-se ao fato de ela ocupar as regiões de clima temperado quase toda Europa, parte da América do Norte, o Japão e a Austrália. Nessas regiões, as quatro estações são bem marcadas, com verões quentes e invernos rigorosos.
As árvores mais comuns são os carvalhos, as castanheiras, as magnólias e as nogueiras. A vegetação herbácea e a arbustiva tendem a ser bem desenvolvidas.
A fauna é diversificada, com animais herbívoros, como esquilos, lebres, coelhos e veados. Os animais carnívoros mais freqüentes são os lobos, linces, texugos e doninhas.
A floresta decídua é ainda muito rica em insetos, pássaros, répteis e anfíbios.
Floresta úmida tropical
A floresta úmida tropical localiza-se entre os trópicos e o Equador. São as florestas de numerosas ilhas do oceano Pacífico, da América do Sul, América Central, África e de regiões da Ásia. Exemplo: floresta Amazônica.
Com vegetação exuberante e grande diversidade de espécies tanto vegetais quanto animais, esta floresta apresenta inúmeras e complexas relações entre os seres vivos.
Nas regiões de florestas úmidas tropicais, o índice de chuvas é alto e as temperaturas são elevadas. Como a vegetação arbórea é muito densa, a luminosidade em seu interior é muito pequena.
A altura das árvores varia muito. As copas das mais altas formam uma camada que recebe toda a luz do sol. Logo abaixo ficam as copas das árvores de menor porte e que recebem os raios solares que conseguiram passar pelas mais altas. Em seguida vêm as copas do arbustos. A medida que nos aproximamos do solo, a quantidade de luz torna-se escassa, mas a vegetação rasteira é geralmente variada.
Entre as espécies vegetais existentes nessas matas, encontramos os ipês, as seringueiras, os jacarandás, os jatobás, os guapuruvus e as canelas. Nos troncos das árvores desenvolve-se grande número de trepadeiras e epífitas, como orquídeas, bromélias e samambaias, além de musgos e liquens.
A fauna é muito rica e variada. Muitos animais são arborícolas, ou seja, vivem nos galhos das árvores. É o caso de macacos, preguiças, lagartos, cobras, pererecas, roedores e morcegos, além de vários pássaros como papagaios, araras, beija-flores, pica-paus, etc.
As populações de insetos são muito variadas: mosquitos, formigas, borboletas, etc. No solo vivem muitos seres, como porcos-do-mato, as antas, além de aves, répteis, moluscos, vermes, aracnídeos e decompositores, como bactérias e fungos. Estes últimos seres têm um papel fundamental na vida dessa floresta. Eles decompõem folhas e galhos que caem das árvores, assim como os restos animais que vão se depositando no chão, transformando-os em matéria inorgânica, que é devolvida ao solo. Assim reinicia-se o ciclo da matéria nesse complexo e surpreendente ecossistema.
Campos, ecossistemas em que as gramíneas predominam
Os campos são ecossistemas formados por uma vegetação predominantemente rasteira, constituída basicamente de gramíneas. Podem também abrigar vegetais arbustivos e arbóreos.
Os campos recebem várias denominações, como cerrado (Goiás, Mato Grosso, Mato Grosso do Sul, Minas Gerais, São Paulo, etc.), estepe (Rússia), pradaria (Estados Unidos), savana (África), etc.
Alguns campos são “limpos”, isto é, possuem uma forte predominância de gramíneas; é o caso dos pampas sulinos, que oferecem excelentes condições para a criação de gado. Outros campos, como os cerrados brasileiros e as savanas africanas, são “sujos”, isto é, além de gramíneas, abrigam inúmeras árvores espaçadas e um considerável número de plantas herbáceas.
A fauna dos campos é variável, conforme o tipo considerado. De maneira geral, abrigam mamíferos de alta velocidade: leopardos, antílopes, zebras e girafas, nas savanas africanas; antílopes, búfalos e coiotes, nas pradarias americanas. Além dos mamíferos, a fauna dos campos inclui aves diversas (gaviões, corujas, etc.), inúmeros répteis, insetos, etc.
A sobrevivência nos desertos
Os desertos são encontrados na África, na Ásia, na Austrália do Norte e na América do Sul. O maior deles é o deserto do Saara (África).
Nos desertos, o solo é árido e as chuvas são muito escassas. O grande fator ambiental que limita a vida animal e vegetal nesses ecossistemas é a água. A vegetação é pobre e pode ser formada principalmente de cactáceas.
A fauna dos desertos varia de um tipo para outro. De maneira geral, é constituída de insetos (grilos, besouros, cupins-de-areia, etc.), répteis (lagartos e cobras) e mamíferos (ratos, gazelas, raposas, coiotes, camelos, etc.), entre outros animais.
Durante o dia, a temperatura desses ambientes pode chegar a 50 ºC. Essas regiões podem ficar sem chuva durante anos. No deserto do Saara, por exemplo, há registros de períodos sem chuva por mais de dez anos em determinados locais.
Veja alguns exemplos de adaptações para a vida nos desertos:
As plantas suculentas, como os cactos, armazenam água no caule e suas raízes são muito espalhadas permitindo a exploração de uma área maior do solo. Muitos vegetais que se desenvolvem nessa região possuem ramos verdes ou folhas minúsculas que caem quando a água começa a faltar, evitando a evaporação. Além disso, suas folhas possuem uma espessa cutícula que reduz as perdas de água por transpiração.
Os animais, por sua vez, alimentam-se de plantas suculentas nas quais encontram, ao mesmo tempo, nutrientes e água.
Muitos animais enterram-se na areia durante o dia refugiando-se do calor excessivo, pois a dez ou vinte centímetros de profundidade, a temperatura da areia se reduz a menos da metade da temperatura da superfície. Muitos animais têm apenas hábitos noturnos. Outras características que permitem a vida dos animais no deserto incluem a formação de urina e de fezes concentradas, a escassez ou ausência de glândulas sudoríparas, etc.
Tundra, um ecossistema muito frio
No pólo norte e seus arredores há apenas duas estações durante o ano: um longo inverno, que dura cerca de nove meses, e um curto verão, com duração de cerca de três meses.
Essa região dispõe de pouca luminosidade, fato que limita o desenvolvimento de uma vegetação exuberante. No verão, desenvolve-se uma vegetação rasteira, composta predominantemente por musgos e liquens, que recebe o nome de tundra.
Essas plantas servem de alimento a animais herbívoros como a rena, o boi almiscarado, os lemingues e as lebres árticas, que, por sua vez, nutrem carnívoros como o lobo ártico, o urso polar, a raposa-ártica e a coruja-das-neves.
No inverno, a vegetação praticamente desaparece, o que provoca a migração de muitos animais para outras áreas, em busca de melhores condições de vida. É o caso, por exemplo, da raposa e da coruja-das-neves, que procuram regiões mais quentes, onde é mais fácil encontrar pequenos animais que lhes sirvam de alimento.
Mas os lemingues permanecem na tundra, mesmo no inverno rigoroso. Esses pequenos roedores não hibernam nem armazenam reservas alimentares para o inverno. Eles vivem em galerias que cavam no gelo e alimentam-se dos tipos de liquens, musgos e outros vegetais que conseguem sobreviver no inverno.